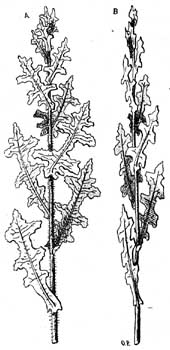
Рис. 31. Lactuca Scariola. A - растение, наблюдаемое с широкой стороны; плоскость расположения листьев есть плоскость меридиана. В - растение, наблюдаемое сбоку; плоскость меридиана расположена косвенно по отношению к плоскости бумаги.
| Сообщества ксерофитов | Е. Варминг |
Для уменьшения испарения в критическое сухое время года или для предотвращения последствий внезапнаго увеличения испарений применяются следующия средства:
1. Периодическое уменьшение поверхности.
2. Сила света, действующая на усвояющие органы, и испарение умеряются профильным положением органов (движения, зависящия от силы света, "фотометрическия движения").
3. Продолжительная установка в профиль (растения-компасы и т. п.; рис. 31).
4. Своеобразныя формы листьев и побегов с незначительной поверхностью.
5. Кроющие волоски, защитные листья и т. п.
6. Анатомическия приспособления для защиты от сильнаго согревания и сильнаго испарения.
Мы разсмотрим в отдельности все эти приспособления, которыя, конечно, свойственны преимущественно многолетним видам.
1. Периодическое уиеньшение поверхности. Наилучший способ для растения ограничить свою испаряющую поверхность - это сбросить при наступлении сухого периода все сильно испаряющия части. Это происходить, во-первых, у всех однолетних растений, отмирающих после созревания семян; сиз-мена же все способны противостоять засухе. В связи с этим процент недолговечных (однолетних) видов в пустынях и других подобных местностях очень велик; в течение короткаго дождливаго периода, продолжающагося иногда 1-2 месяца, эти растения совершают свой жизненный цикл - прорастают, цветут, приносят семена и умирают, так что сухое время года они переносят в зародышевом состоянии внутри семян (ср., например, Volkens, II). То же самое происходит у всех луковичных и клубненосных растений, подземные побеги которых в сухое время года содержать запасы питательных веществ и воды; наземные побеги с большими испаряющими поверхностями в сухое время года сбрасываются и жизнь остается скрытой в подземных частях. При наступлении влажнаго периода эти виды торопливо развивают новые наземные побеги и цветки. Путешественники часто упоминают с удивлением о быстром наступлении весны в степях, пустынях и т. под. местностях после первых ливней. В третьих, то же самое происходит у древесных растений, которыя перед наступлением сухого времени года или в течение его, а также зимой сбрасывают листву (листопад). В течение этого времени все наземныя части защищены от испарения пробкой и почечными чешуями, покрытыми пробкой или другими, предохраняющими от испарения образованиями.
У всех этих растений лист, вообще являющийся верным отражением климата, не имеет ксерофильнаго строения или лишь в слабой степени ксерофилен.
Совершенно иначе уменьшается испаряющая поверхность у других растений, например, у некоторых трав, свертывающих в сухую погоду свои листья, так что даже широкие листья становятся трубчатыми или нитевидными. Примером могут служить песчанка (Psamma arenaria), Coryne-phorus canescens, виды Festuca, Anthoxanthum odoratum и многие другие злаки, растущие в степях, на пустошах и на дюнах; в средиземноморских странах, например, виды Stipa, Lygeum, Aristida (Duval-Jouve; Tschirch; Warming, VII, и др.), в Новой Зеландии по Diels'y Poa anceps и Festuca arundinacea. To же явление наблюдается у злаков, растущих на солончаковой почве, например, Triticum junceum. По мере того, как возрастает сухость воздуха, листья свертываются и испаряющая верхняя поверхность, на которой исключительно или преимущественно расположены устьица, оказывается защищенной от испарения, так как устьица таким образом заключаются в безветренныя пространства. В сырую погоду листья снова развертываются. У Cyperaceae мы замечаем подобныя же, только более слабыя движения. У этих последних играют роль сочленовныя клетки (cellules bulliformes, Duval-Jouve), расположенныя в бороздах верхней поверхности листа; эти клетки выше остальных эпидермальных клеток и стенки их, состоящия из клетчатки, сгибаются при заворачивании листа. Движущая сила, по-видимому, заключается прежде всего в лубяной ткани, расположенной обыкновенно на нижней стороне листьев и, смотря по обстоятельствам, то поглощающей воду, то выделяющей ее и сообразно с этим разбухающей или сжимающейся. Однако, тургор мякоти листа также играет, по-видимому, в известных случаях важную роль (Duval-Jouve; Tschirsh, II).
Некоторыя двудольныя растения имеют подобныя же движений, например, Hieracium Pilosella, Antennaria dioica, Crepis tectorum (no WiIIe), вест-индские виды Croton, Euphorbia Paralias (западно и южно-европейское дюнное растете, по Giltay) Листья Erica Tetralix на сырой почве менее свернуты, чем на сухой (Graebner); то же самое можно сказать относительно листьев Ledum palustre. Из тайнобрачных можно назвать упомянутые папоротники и некоторые мхи, а именно виды Racomitnum и Polytrichum; в сухую погоду листья R. canescens сложены и побеги кажутся совершенно серыми от густых длинных волосков; в сырую погоду, а также на сырой почве они звездообразно расправляются. У Polytrichum края листа могут прикрывать ассимилирующия тонкостенныя клетки, расположенныя в средине листа.
2. Движения, которыми регулируется освещение. Некоторыя растения чрезвычайно чувствительны к силе света, а листья или листочки их способны производить движения, регулирующия освещение, причем пластинка листа образует у них при известной степени освещения определенный уголь с падающими лучами; при умеренном освещении (например, в утренние часы) пластинки листа располагаются наиболее выгодно по отношению к свету, чтобы лучи света падали на них под прямым углом (плоскостное расположение). По мере увеличены силы света пластинки располагаются так, чтобы свет падал на них всегда под более острыми углами (профильное положение). Благодаря этому, оне меньше освещаются и нагреваются, и испарение, вследствие этого, также должно уменьшаться. Сюда относятся многия растения со сложными листьями, особенно из числа тропических низких кустарников, например, многие виды Acacia и др. Mimosaceae, многие Papilionaceae и Caesalpinaceae, Oxalidaceae (между прочим, Oxalis Acetosella), Zygophyllaceae. Такия же, зависящия от силы света движения мы встречаем и у растений с простыми листьями, например, у Hura crepitans, Bauhinia. У названных растений листья также не имеют ксерофильнаго строения. Листья, например, вест-индских бобовых растений, обладающие способностью двигаться сообразно с силой света, часто даже и покрыты тонкой и голой кожицей (Warming).
3. Устойчивое положение по отношению к свету у растений - компасов и других растений. Ослабление действия солнечнаго света и вместе с тем уменьшение испарения достигаются еще следующим образом: ассимилирующия поверхности растения в течение продолжительнаго времени занимают профильное или подобное положение, вследствие чего сильные лучи полуденнаго солнца падают на них под острым углом. Профильное положение, которое вышеупомянутый растения принимают во время сильнаго освещения, у этих растений сохраняется после того, как оно было вызвано во время роста и развития растения направляющим действием света. Сюда относятся из представителей нашей флоры Lactuca scariola, листья которой в местностях сильно освещенных располагаются ребром к плоскости (рис. 31; Stahl, I, III). Из других компасных растений упомянем особенно Silphium lacmiatum (Сев. Америка).
Листья, расположенные ребром, встречаются у многих других видов, например, у многих австралийских видов Eucalyptus и Proteaceae, южно-африканских видов Statice, у Conocarpus erecta и др. в Вест-Индии, у Bupleurum verticale (Испания) и т. д. (рис. 32, 33 и 34).
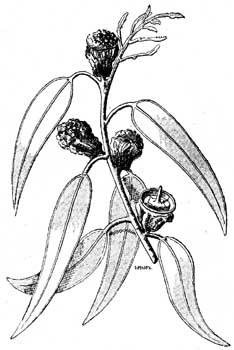
У ксерофитов, растущих на сильном солнечном свету, листовыя пластинки часто стоят перпендикулярно вверх или сильно приподымаются, например, у Coccoloba uvifera и у многих других вест-индских растений, у многих злаков (Brachypodium ramosum, Festuca ovina и др.), Calluna, Рейcedanum Cervaria (no Altenkirch'y), Helichrysum arenarium и др. Реже встречаются свешивающияся или загнутыя вниз листовыя пластинки. Сюда же близко примыкают прутообразные побеги.
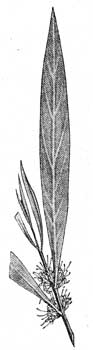
Рис. 33. Вертикально расположенные листья у Hakea saligna (Proteасеае). Австралийская флора. |

Рис. 34. Leptospermum resmiterum (Myrtaceae); вертикальное положение листьев. Австралийская флора. |
Складки и морщины листовых пластинок действуют, вероятно, подобным же образом и оне тем многочисленнее, чем суше климат, например, в Вест-Индии у Lippia involucrata, Plumeria alba и многих др. (Johow, I), в степях Египта у Salvia и Stachys Aegyptiaca, Pulicaria и Urginea undulata и др. (Volkens); в Дании, например у Vicia Cracca (Warming, также Heinricher, 1) Складки листьев пальм, Cecropia и других относятся, вероятно, тоже сюда. Солнечные листья в этом отношении отличаются от листьев теневых (по Johov'y).
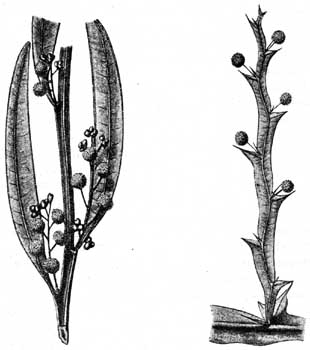
Так как эти различные типы расположения растительных органов достигаются обыкновенно лишь во время развития индивидуума, благодаря поворотам, изгибам, то, вследствие этого, листья у всех видов из числа названных растительных форм, смотря по природе местности, будут устроены различно. Листья, подверженные солнечному свету, засухе и ветру, будут гораздо более подняты вверх, расположены ребром, более складчаты, чем листья, находящиеся в тени и в сырости, особенно в сыром воздухе; это показывают, например, Calluna, Juniperus communis, Lycopodium Selago и alpinum (изображения у Warming, V).
Следующия растения имеют прирожденное профильное расположение: австралийския акации с филлодиями (листовидными, но отвесно стоящими листовыми черешками, лишенными листовых пластинок), а также многия растения с плоскими или крылатыми стеблями (рис. 34), или с нисходящими листьями, например, Baccharis triptera в Бразилии, Genista sa-gittalis, Muehlenbeckia platyclada, Carraichaelia australis, виды Colletia и др. Эти виды побегов в то же время почти безлистны; стебель заменяет собой листья. Сюда же принадлежит мечевидный лист у Iridaceae, Tofieldia, Narthecium и Sisyrynchium.
4. Формы листьев и побегов с незначительной поверхностью. У многих ксерофитов испаряющие органы, именно листья, чрезвычайно сокращаются в своей величине; вследствие этого, получаются отступления от обычной физиономии всего побега, целый ряд особых ксерофитных форм побегов в различных последовательностях. Недостаток воды вызывает уменьшение поверхности: один и тот же вид на сухой почве имеет мелкие листья, на сырой большие, например, Urtica dioica, Viola canina, Erodium cicutarium; многия пустынныя растения развивают при наступлении дождливаго времени года большие листья впоследствии же являются более мелкие листья или они почти совсем отсутствуют, например, у Zilla, Alhagi и др. Величина листа является прямым следствием освещения и сухости (ср. Henslow, Elliot, Groom и др.). Недостаток воды способствовал вероятно, образованию ряда определенных типов растений, характеризующихся относительно незначительной ассимилирующей работой, следствием чего является замедленный рост.
А. Формы листа.
1. Игловидный или пиноидный лист, похожий на хвои (у хвойных Proteaceae, Ulex Europaeus и у др. вечно-зеленых деревянистых растений). Он имеет длинную, линейную, острую форму (рис. 35.) и хлорофильная ткань расположена более или менее в центре; ложбинок или кроющих волосков не бывает.
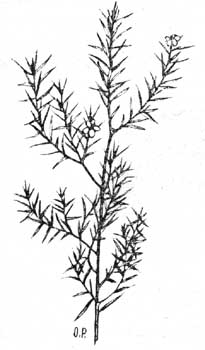
Рис. 35. Ветка Coleonema album. Пример игловидных листьев из капландской жестколистной флоры. |
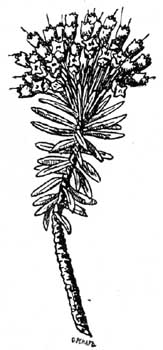
Рис. 36. Erica multiflora. Пример эрикоидных листьев из средиземноморской жестколистной флоры. |
Зависимость такой формы листа от испарения выражается в том, что поверхность листа по отношению к объему его гораздо меньше, чем у плоскаго листа с тем же объемом, и, вследствие этого, испаряющая поверхность меньше. То же самое можно сказать и относительно следующих листовых форм.
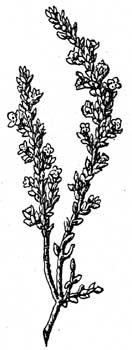
2. Эрикоидный лист. Это завернутый лист, края котораго свернуты вверх или вниз (первое реже, например, у Passerina); получается, таким образом, безветренная, обыкновенно еще покрытая волосками бороздка (рис. 25, 36 и 37), в которой скрыты устьица; устьица обыкновенно приподняты над уровнем эпидермиса, совершенно также, как у шерстисто-волосистых листьев. Эрикоидные листья часто коротки, линейны и встречаются у многих вечнозеленых растений, как Erica, Calluna, Cassiope и др. Ericaceae, E.mpetrum, Epacridaceae, Proteaceae, Myrtaceae, Berberis empetrifolia (Чили), у южно-африканских Thymeleaceae, Compositae, Rubiaceae и видов других семейств, растущих в местностях, где сильное испарение. У Passerina (рис. 38), Helichrysum depressum (no Diels) безветренная ложбинка листа обращена к стеблю и лист к нему прижат.
3) Чешуйчатый лист, широкий и короткий, направленный вверх, прижатый, иногда, у кипарисовидных побегов, сбегающий, встречается у многих Cupressaceae и растений самых разнообразных семейств, например, у Scrophulariaceae (Veronica thuyoides и cupressoides на горах Новой Зеландии), Santalaceae, Tamaricaceae, Francoaceae, Compositae, Umbelliferae (например, Azorella на высоких горах Южной Америки и в антарктических областях). Такие побеги Goebel относит к особому типу - Lepidophyllum.
4) Щетиновидный или нитевидный лист встречается у многих травянистых, злакоподобных однодольных; верхняя поверхность такого листа большей частью покрыта бороздами или имеет вид желоба, устьица спрятаны в бороздах, усаженных волосками (рисунки у Kerner, H). Складывание, зависящее от степени сырости, встречается часто, например, у Festuca ovina, Corynephorus canescens, у многих злаков степей и у злаков пустынь. Разсе-ченные листья представляют такия же, очень небольшия цилиндрическия лопасти (например, у Artemisia campestris).
5. Сюда же примыкает ситниковидный (юнкоидный) лист; это длинный, лишенный бороздок, цилиндрически лист (виды Juncus, многия Cyperaceae, некоторыя Umbelliferae в высоких горах Южной Америки, Goebel, II, 2 ч.). Эта форма встречается преимущественно на сырой холодной почве, открытой доступу ветра.
6. Здесь можно упомянуть о форме листа у сочных растений (Succulentae), так как здесь лист часто является, независимо от толщины, более или менее цилиндрическим, линейным, удлиненным или лопатковидным, не имеет ни зубчиков, ни других надрезов (например, Sedum acre, Sempervivum tectorum, виды Mesembrianthemum, Orchidaceae); эта форма представляет меньшую испаряющую поверхность, чем в том случае, если бы эта же масса занимала большую площадь. Мнение Henslow'a, что сочность растения прямо обусловливается действием окружающей среды, вероятно, правильно.
7. Встречается еще много других форм листьев, которыя не могут быть отнесены ни к одному из числа вышеуказанных листьев; они представляют различныя приспособления, обусловливаемый испарением; некоторые листья велики, кожисты (например, Loiseleuria procumbens, Phillyrea; рис. 39). Другие узки и тверды, более или менеесвернуты, например, у Lavandula (рис. 40) и Hyssopus и др. видов в средиземноморских странах; друтие шире и более плоски (Dryas, Rhododendron Lapponicum и др.). Подобные листья обыкновенно защищены от испарения другими средствами, о которых речь впереди (относящаяся сюда литература у Vesque, Volkens, Goebel, Warming, Henslow).

Рис. 39. Phillyrea media. Средиземноморская жестколистная флора. |

Рис. 40. Lavandula Stoechas. Средиземноморская жестколистная флора. |
Побеги растений с вышеуказанными листовыми формами (особенно 1, 2 и 3) обыкновенно обильно покрыты листьями. Незначительная величина листьев возмещается количеством их; кроме того, листья скучены на коротких побегах, и это обстоятельство, по всей вероятности, ослабляет испарение.
В. Формы побегов.
"Безлистные" побеги, т. е. побеги, снабженные сильно редуцированными или скоро опадающими листьями, встречаются у многих ксерофитов; обыкновенный лист исчезает и функцию его исполняет стебель, вырабатывающий палиссадную ткань. Эти побеги представляют следующие типы.
1. Крылатые, часто безлистные побеги (рис. 34).
2. Прутевидные побеги (форма Spartium). Побеги имеют вид прутьев, прямы, стройны, часто сильно ветвятся; листья у некоторых видов еще довольно велики (например у Genista tinctoria, Spartium junceum), но обыкновенно скоро опадают; у других же растений они с самаго начала редуцированы и лишены всякой функции. Во всяком случае листья далеко отстоят друг от друга, так что стебель может быть освещен сбоку. Стебли круглы, часто покрыты глубокими бороздами, на дне которых помещаются устьица и палиссадная ткань, между тем как ребра содержат механическую ткань. Эта форма очень распространена среди некоторых Leguminosae, свойственных средиземноморским странам (особенно среди Genisteae: виды Genista, Retama, Cytisus), Casuarina, Ephedra, многих Chenopodiaceae, например у Anabasis (которая, однако, тесно примыкает к солончаковым растениям), у Capparis aphylla, Periploca aphylla, Polygonum equisetiforme и др. (Литература: Pick, Volkens, Schube, Ross, Nilsson, Briquet).
3. (Ситниковидный) юнкоидный побег. У многих видов Juncus и у Сурегасеае встречаются высокие, округлые, безлистные и неразветвленные побеги (по форме они сходны с листьями некоторых из этих видов). Между объемом и поверхностью здесь будет тоже отношение, как это приведено выше. Такой же формы побеги встречаются у многих болотных видов этих двух семейств (Scirpus lacustris, Heleocharis palustris, у Junci genuini и т. д.
Сюда же относятся, например Restionaceae.
4. Иглообразный кладодий у Asparagus.
5. Плоский побег; безлистный плоский побег, с ветвями, обыкновенно располагающимися вертикально или профильно; у Muehlenbeckia platyclada (Coccolola p.), Ruscus, Semele, Phyllocladus, Carmichaelia australis и др., Viscum и видов Phoradendron (рис. см. особенно у Kerner'a, I).
6. Шиповидный побег, например у Colletia. 7. Кактусовидная форма с различными подразделениями у Cactaceae, Euphorbia, Stapelia; она будет еще раз упомянута при описании сочных растений.
В заключение следует заметить, что многия из указанных форм листьев и побегов снова встречаются среди солончаковых растений.
5. Уменьшение испарения посредством кроющих органов. Если испаряющая поверхность будет покрыта воздухоносными органами и воздух как в этих органах, так и между ними будет более и менее неподвижен, то, понятно, испарение будет сильно уменьшено. Это средство применяется многими ксерофитами различным образом.
а. Кроющие волоски. Здесь с особенной очевидностью обнаруживается разница между гидрофитами и ксерофитами; первые почти всегда голы, вторые часто сильно серо - или бело-войлочны и шерстисты, или серебристы; такия окраски обусловливаются содержанием воздуха в волосках и в промежутках между ними; если все волоски направлены в одну сторону, тогда получается серебристо-блестящая поверхность. Кроющими волосками могут быть только отмершие, наполненные воздухом, следовательно сухие волоски. Уже давно известно, что виды, обыкновенно голые, в сухих местах покрываются волосками, а покрытые волосками, делаются здесь еще более волосистыми, чем в сырых местах (Ranunculus bulbosus, Polygonum Persicaria, Mentha arvensis, Stachys palustris и т. д.) Этиолированные побеги картофеля в сыром воздухе почти голы, в сухом - покрыты волосками (Vesque). Многочисленныя растения, встречающияся на скалах Средиземнаго моря или среди сухих кустарников Вест-Индии, разнообразным степныя, пустынныя, а также горныя растения покрыты пушистыми волосками. Пожалуй наиболее волосистым является одно сложноцветное растение высоких гор Южной Америки - Espeletia из области Парамосов в Венецуеле(рис. 41) по Goebel, II. Волоски представляют из себя экран, защищают растение от солнца, смягчают внезапныя колебания температуры и подобно войлоку задерживают испарение. Особый вид кроющих волосков представляют щитовидные волоски, придающие покрытому ими растению своеобразный серебристый блеск, например у Elaeagnaceae, видов Croton и др. Кроющие волоски почти всегда гуще всего располагаются на нижней стороне, где находятся устьица. Эти волоски, находящиеся на затененной стороне, конечно, не могут иметь иного назначения кроме ослабления перемены воздуха. Молодые стебли и листья часто бывают гуще покрыты волосками, чем старыя части, что стоит в связи с большей потребностью их в защите от сильнаго испарения. В сухих местностях тропических стран первые листья, развивающиеся после сухого времени года, бывают иногда более пушисты и, вообще, имеют совершенно другой вид, чем листья, появляющиеся позже; эти последние больше и зеленее (H. Schinz). Нужно, однако, заметить, что есть одна группа ксерофитов, а именно сочныя растения, которыя совсем не имеют кроющих волосков и обыкновенно совершенно голы (виды кактусов, Aloe, Agave и др.).
Образование волосков является несомненно, подобно всем другим средствам самоприспособляемости (саморегулирования) растений, прямым приспособлением к климату. Относительно действующих причин Henslow (I), опираясь на замечание Mer'а, высказывает мнение, что волоски появились благодаря местному усилению питания в связи с угнетением паренхимы: по мере угнетения паренхимы, как компенсация ея развивались волоски. С этой гипотезой мы, однако, не далеко уйдем вперед в понимании соотношения (корреляции) между развитием волосков и засухой, если только она вообще верна.
в. Покрывающие листья служат для защиты молодых частей побегов от сильнаго испарения и интенсивнаго света. Сюда принадлежат, во-первых, почечныя чешуйки, в типическом виде находящияся у меняющих листья деревьев умереннаго и холоднаго климатов, но встречающияся также и в тропических странах. Назначение этих чешуек - защищать покоющиеся в почке молодые листочки от испарения и предохранять почки во время развития листвы от перемен температуры, что они могут исполнять благодаря опробковению, образованию волосков, выде-лению смолы и т. д. (Griiss, Feist, Cadura и др.).
Прилистники и листовыя влагалища (например у некоторых дюнных злаков) могут служить для той же цели, хотя их и нельзя называть почечными чешуями в узком смысле слова, точно также как и пленчатые прилистники видов Paronychia, Herniaria и др., одевающие часто молодыя части побегов плотным, серебристым покровом. Старые листья и остатки листьев во многих случаях служат для той же цели. "Влагалищными злаками" (Tunicagraeser) Hackel называет такия травы, у которых нижния части листьев после отмирания верхних частей сохраняются или в виде плотных замкнутых влагалищ, или в виде мочалистых нитей. Такие остатки листьев встречаются у трав, растущих на дюнах, в степях и пустынях (например у Nardus stricta, Andropogon villosus, Scirpus paradoxus, S. Warmingii, видов Aristida); роль их состоит в том, что они уменьшают испарение и задерживают воду (Hackel; ср. также Warming, XI; Henslow). To же самое встречается у Velloziaceae, растущих на вершинах гор и на высоких равнинах Бразилии (Warming, XI). У некоторых видов Oxalis, преимущественно у южно-африканских, встречаются своеобразно развитые кроющие листья на луковицах (Hildebrandt). Здесь же следует упомянуть о плотных дерновинах, состоящих из тесно сплоченных побегов и их остатков, встречающихся всюду в приледниковой растительности, особенно южно-африканской; оне обыкновенно бывают так тверды, что их едва можно разрезать или разрубить; тут один побег защищает другой, старыя части защищают молодыя.
Корни некоторых эпифитных растений защищены от испарения посредством листьев, которые плотно налегают на них и задерживают сырой воздух, например у Conchopyllum imbicatum, Dischidia, Pothos (Goebel II, и часть; рис. 10, стр. 128, и 42).

Корни некоторых египетских пустынных злаков (виды Aristida, Andropogon, Elionurus, Panicum, Sporolobus) заключены в песчаныя трубочки, состоящия из песчинок склеенных особым клейким веществом, выделенным корневыми волосками. Volkens видит в этом, защиту от испарения. Однако возникает сомнение, имеют ли эти корневые покровы какое-либо особое назначение или являются прямым следствием дыхания корней и выделения ими угольной кислоты; таким образом часто возникают на корнях инкрустации углекислаго кальция.
6. Анатоническия и другия особенности строения, уменьшающия испарение. И в этом отношении существует резкое различие между ксерофитами и гидрофитами. Особенно различаются по своему строению у этих растений следующия ткани.
А. покровныя ткани,
В. хлорофиллоносная ткань,
С. система проветривания, а именно: а. устьица и в. межклетныя пространства.
А. Покровныя ткани. Очевидно, что кожица, постоянно окруженная водой или сырым воздухом, и кожица, окруженная сухим воздухом, подверженная сильному испарению, должны представлять большия различия.
Эпидермис. Кутикула служить важным регулятором испарения; толщина ея соответствует потребности растения ограничить испарение. Кутикула у гидрофитов обыкновенно очень тонкая, у ксерофитов большей частью толстая. Наружныя стенки эпидермальных клеток сильно утолщены и кутикуляризованы; в некоторых случаях отлагаются даже кристаллы щавелевокислаго кальция или кремнекислоты. Эпидермальныя клетки приобретают многогранную форму и стенки их прямыя. Вследствие свойств эпидермиса листья становятся кожистыми, что предохраняет их от скручивания, и блестящими, что составляет общую, бросающуюся в глаза особенность тропических деревьев, встречающуюся также и у растений умереннаго климата с многолетними листьями (например у Ilex Aquifolium, у многих хвойных, Vinca и др.). Этот блеск доказывает, что часть световых лучей отражается листьями, что, может быть, полезно для них (Wiesner, I). Кутикула часто бывает снабжена тонкими перекладинами, особенно в тех случаях, когда клеточная стенка имеет вид свода. Vesque (II) и Henslow(I и II) высказали гипотезу, по которой вышеупомянутое обстоятельство служит для разсеивания и ослабления лучей света.
Воск, выделяющийся на поверхности у многих ксерофитов уменьшает испарение, что Tschirch и Haberlandt доказали путем опытов. Например, у Capparis spinosa растущей в египетской пустыни, в начале сухого времени года образуется на листьях толстый слой воска, который без сомнения совершенно уничтожает испарение (Volkens). Слои воска могут быть очень толстыми, например у Sarcocaulon (Южная Америка) толщина воска больше 1 mm.
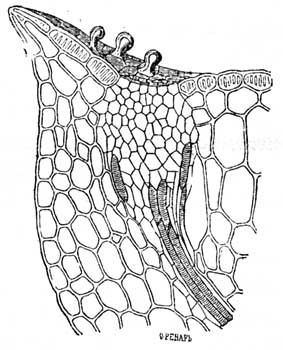
Листья с сизым налетом не имеют обыкновенно острых зубчиков по краю листа, самое большее они имеют - зубчики закругленные и снабженные аппаратом выделяющим воду.
На поверхности некоторых пустынных растений образуется кора из солей, которая днем придает им серую окраску и защищает от испарения; к ночи этот покров расплывается, так как поглащает влагу из атмосферы. Железки некоторых Plumbaginaceae и видов Saxifraga, выделяющия известь, также препятствуют испарению (рис. 43 и 44). "Лакированные" листья. Смола или тому подобный вещества выделяются волосками на поверхности листьев многих ксерофитов, особенно в южном полушарии. Листья вследствие этого становятся клейкими и "лакированными", на их поверхности выделяется блестящий, стекловидный слой; стенки эпидермиса тонки и слегка кутикуляризованы (Volkens).
Слизь в некоторых случаях выделяется волосками (коллатеры Ганштейна); например, в почках Polygonaсеае, она может быть служит для принятия воды и, вероятно, умеряет испарение. Предположение, что слизь, выделяющаяся на поверхности лакированных листьев, служит для поглощения воды, совсем еще не доказано (Mei gen, II).
Содержимое эпидермалных клеток также может служить приспособлением для уменьшения испарения. Уже безцветный эпидермис является водохранилищем (Westermaier); в связи с этим листья подводных частей гидрофитов покрыты зеленым эпидермисом. Присутствие некоторых веществ в содержимом эпидермиса делает его менее проницаемым для водяных паров.
Неизвестно, играет ли какую-нибудь роль дубильное вещество, встречающееся часто в эпидермисе, особенно зимою у зимующих листьев (Warming IV), а также у степных и пустынных растений, например у Alhagi, Monsonia, Astragalus, Tamarix, где оно, по-видимому, находится в связи с водоносной тканью (Volkens, Henslow, I). Наоборот, антокиан - красное вещество, встречающееся у многих растений, преимущественно в эпидермисе, имеет то значение, что поглащает часть световых лучей - именно так называемые химические лучи. Известно, что антокиан часто встречается на молодых частях растений и в проростках, нуждающихся в защите от сильнаго света и связаннаго с ним сильнаго испарения (особенно под тропиками; при развитии листвы многие молодые побеги являются красно-бурыми; Pick, Kny).
В заключение следует упомянуть об ослизнении внутренних стенок эпидермиса. У многих ксерофитов, особенно у древесных растений, эти стенки ослизняются (например, у Empetrum, Arbutus Unedo и др. Ericaceae). Быть может это служит для уменьшения испарения (Volkens), быть может таким образом получается водохранилище; ср. стр. 261 (Westermaier, Tschirch, см. также Walliczek, Vesque, Radlkofer, Gruber).
Пробка. Изледования и опыты ясно обнаружили, что пробковая ткань может уменьшать испарение благодаря существованию воздуха в клетках и другим своим качествам. Толщина пробки иногда находится в прямой зависимости от сухости климата, что обнаруживается, например в различии между деревьями бразильских кампосов и прилегающих к ним лесов. Изсушающее влияние пожаров в кампосах, повидимому, способствует дальнейшему росту пробковой ткани; это одно из явлений саморегулирования природы (Warming, VIII). Толстые пробковые покровы встречаются у многих пустынных растений, например, у Testudmaria Elephantipes в южной Африке, Cocculus Leaeba в Египте.
Наконец, здесь следует еще указать на факт, имеющий, может быть, наибольшее значение и служащий главной причиной указанных выше особенностей строения, поскольку оне касаются эпидермиса: части растения, смачиваемыя водой легче вянут, чем те, которыя не могут быть смачиваемы. Причину такого усиленнаго испарения Wiesner видит в своеобразном набухании клеточных стенок от воды; этим уменьшается способность клеточных стенок не пропускать воду. Многия указанныя средства, уменьшающия испарение, защищают также части растений от смачивания водой и этим ослабляют сильное испарение.
В. Хлорофиллоносная ткань. Характерной чертой ксерофитов является значительное развитие палиссадной ткани, причем или увеличивается число клеточных слоев, или возрастает высота слоев (длина клеток), или то и другое вместе. На стр. 23 упоминалось разногласии во мнениях, стремящихся объяснить эти особенности строения, и было высказано предположение, что это обстоятельство находится в ближайшей связи с сухостью воздуха и сиспарениеы. Клетки паллисадной паренхимы в упомянутых на стр. 241 вертикальных органах направлены косо н поверхности, совершенно также как и в безлистных стеблях.
С. Система проветривания. Межклеточныя пространства представляют те места, где испаряется вода, и так как испаряющая поверхность растения измеряется не только поверхностью, непосредственно граничащей с атмосферой, но также поверхностью стенок всех межклеточных пространств, то можно, a priori сказать, что межклетныя пространства, наполненныя воздухом у ксерофитов будут значительно уже, чем у гидрофитов, где они очень велики, как это приведено на стр. 158 (исключение - гидрофиты литофильные). На ряду с этим распределение и число устьиц в обеих группах растений представляют большое различие.
a. Schwendener нашел, что устьица по своему строению приспособлены к регулированию испарения; они замыкаются, когда есть опасность сильнаго испарения и открываются, когда испарение прекращается. Замыкающия клетки некоторых растений, свойственных пустыням, обладают подвижностью только в молодых листьях; в старых листьях, благодаря сильным утолщениям стенок, эти клетки неподвижны, и щель может быть закупорена воском и смолой (Volkens, GiIg). Stahl (VI) опубликовал несколько важных изследований относительно функций устьиц, из которых между прочим видно, что у болотных и солончаковых растений устьица не могут замыкаться, что, однако, оспаривается другими.
Число устьиц зависит от характера окружающей среды. Нормально, погруженныя в воду части гидрофитов обыкновенно лишены устьиц, которыя здесь были бы безполезны; что касается наземных частей растений, окруженных воздухом, то можно в общем принять за правило, что чем суше местообитание, тем меньше число устьиц, что лучше всего видно при сравнении близко родственных видов (Pfitzer, Zingeler, Czech, Tschirch, Volkens, Altenkirch). Распределение устьиц находится в тесной зависимости от испарения и условий влажности. У луговых злаков обыкновенно обе поверхности листа снабжены устьицами, у степных же обыкновенно только верхняя, покрытая бороздками, поверхность (Pfitzer); у других ксерофитов устьица обыкновенно расположены только на нижней поверхности, где они часто спрятаны под образованиями, затрудняющими испарение.
У некоторых ксерофитов устьица погружены ниже уровня поверхности в углублениях, бороздках и т. д., защищенных часто волосками, вследствие чего воздух с трудом выходит из устьиц и, следовательно, уменьшается испарение. Погружение происходит следующими способами (ср. Tschirch, I):
В простейшем случае вокруг отдельных устьиц образуется чашевидное или воронкообразное углубление, или вследствие того, что кутикула выдается вперед в виде каймы (передний дворик), или потому, что соседния клетки подымаются выше устьиц, лежащих ниже уровня поверхности листа (наружная дыхательная полость, Tschirch, I), например у Pinus silvestris, у Proteaceae и др. У Euphorbia Paralias устьица окружены низкими сосочками (Giltay). У Nerium, Banksia и др. устьица собраны группами в углублениях, находящихся на нижней поверхности листьев; вход в эти углубления сужен и более или менее закрыт волосками. У очень многих растений устьица расположены в продольных бороздках, причем обыкновенно в тех, края которых усажены более или менее густо волосками. Многие стебли, особенно имеющие прутовидную форму, снабжены глубокими бороздками, в которых только и находятся устьица (Casuarina, Ephedra, Acanthosicyos horrida, виды Genista и т. д.). Борозды на верхней поверхности листьев имеютсяу многих степных злаков; здесь эти бороздки часто кверху еще сужены, и устьица вследствие свертывания листьев могут быть еще более скрытыми (Corynephorus canescens, Festuca ovina, Aristida, Stipa, Sporobolus spicatus, Cynodon Dactylon и т. д.); ср. также стр. 230. Ha нижней стороне листьев многих ксерофитов находятся бороздки, усаженныя волосками, или более широкие желобки, например у Empetrum (Griiber), Phyllodoce coerulea, Calluna, видов Erica, Loiseleuria procumbens, Ledura palustre, Cassiope tetragona (Warming, V), Dille nасеае (ср. Steppuhn в Bot. Centralbl., LX II) и т. д. (рис. 43). Сюда же можно причислить те листья, края которых менее свернуты и устьица которых лежат на нижней поверхности, усаженной волосками, например листья Dryas octopetala. Если листья постоянно приподняты кверху и прижаты к стеблю, так что их нижняя сторона наиболее освещена солнцем, то эта последняя может получить характер, свойственный верхней стороне листа; именно на ней может развиться палиссадная ткань; бороздка с устьицами находится в таком случае на верхней стороне листа (например у Passerina fiiliformis, по Goebel'ю, II, у Lepidophyllum quadrangulare, Phoenocoma prolifera). Таким образом, такия растения имеют несколько средств для задерживания водяных паров.
Что эти обстоятельства находятся в прямой зависимости от сухости климата, это доказывается такими видами как Ledum palustre и Andromeda Polifolia; чем больше эти растения подвержены ветру и сухости, тем меньше у них листья и тем более свернуты края их.
В последних приведенных случаях мы видим переход к плоским и широким листьям, нижняя поверхность которых густо покрыта войлочными, щитовидными или другими кроющими волосками, служащими единственной защитой или прикрытием устьиц. Такие листья представляют иногда на своей нижней поверхности выпуклую нервацию; в петлях сети жилок лежат устьица, следовательно, все же немного погруженный (например у Lantana involucrata в Вест-Индии).
Если устьица лежат в пространствах, защищенных от ветра и наполненных водяными парами или густо покрытых волосками, то они приподняты над поверхностью листа, подобно тому, как это имеет место у листьев, живущих в сыром воздухе.
b. Межклетныя пространства. Уже строение дыхательных полостей может служить для регулирования испарения, например тем, что стенки их становятся кутикуляризованными, или тем, что оне окружаются особыми клетками (Restionaceae), или, наконец, тем, что делаются очень маленькими. В некоторых случаях кутикула с наружной поверхности эпидермиса переходит через устьица на стенки дыхательной полости (de-Bary, Vergleichende Anatomie, S. 79).
Вообще, вследствие вышеприведенных причин, межклетныя пространства у ксерофитов очень узки (ср., например измерения Altenkirch'a относительно дыхательных полостей). Однако встречаются исключения, например у Restionaceae, у которых кроме узких кольцевых каналов есть большия воздушныя пространства, играющия, быть может, известную роль в ассимиляции угольной кислоты; также велики они у многих свернутых листьев, например, у Empetrum и, по Volkens'y, у различных пустынных растений, превосходно, однако, защищенных от сильнаго испарения. Но межклетники без сомнения помогают также и ассимиляции и там, где лист достаточно защищен от испарения другими приспособлениями, там они могут быть сполна приспособлены к исполнение именно этой задачи.
Упомянутые узкие каналы встречаются также у Hakea suaveolens (австралийское пустынное растение), Olea Europaea, Kingia (Tschirch), у некоторых песчаных злаков (Festuca rubra и Triticum acutum; Giltay) и у других растений; это узкия межклетныя пространства, которыя в виде лент проходят поперек клеток столбчатой паренхимы; эти обходы, конечно, должны задерживать выход воды. Некоторыя растения пустынь, например Cynodon Dactylon и Sporobolus spicatus имеют сплетение из чрезвычайно тонких, перепутанных межклетных каналов (Volkens); однако нельзя с уверенностью сказать, служат ли все эти различныя формы межклетных пространств для уменьшения испарения.
7. Другия средства регулирования испарения. Эфирныя масла встречаются преимущественно у ксерофитов. Гариги и маккия (маквисы) Средиземноморских стран, кампосы Бразилии и другия сообщества благоухают от Cistus, Labiatae, Verbenaceae, Compositae, Myrtaceae, подобно тому как наши песчаныя поля благоухают тимьяном, а степи Азии полынками. Причинная зависимость между сухостью климата и почвы и появлением масла не выяснена, неизвестна также и польза, приносимая им. Эфирныя масла испаряются легче, чем вода и окружают растете благоухающей атмосферой По Тиндалю воздух, богатый эфирными маслами, гораздо менее диатермичен, т, е. он гораздо менее способен пропускать лучистую теплоту, чем обыкновенный воздух; вследствие этого эфирныя масла, может быть, способны днем умерять освещение, а через это и испарение, ночью же, при чистом небе, благодаря им будет ослаблено охлаждение. Возможно также, что эфирныя масла приносят и другую пользу, например служат защитой от травоядных животных, на что особенно указываешь Stahl.
Значение млечнаго сока тоже не вполне выяснено; по мнению одних - млечные сосуды являются проводящими путями (Haberlandt, Schullerus, Pirotta и др.), по мнению других (например Kerner) - они служат защитой от животных.